
超分子是指由两个或多个分子通过非共价分子间相互作用结合在一起的有序分子聚集体。超分子作用广泛存在于自然界,在生物传感、分子成像、生化过程等研究领域发挥着重要作用。目前,可采用核磁共振谱、紫外-可见吸收光谱、荧光光谱等技术对超分子聚集体中的非共价相互作用进行表征。然而,上述技术虽然可以表征大量分子的平均行为特征并给出详细结构信息,但是无法满足表征单个超分子组装体微观动态特征、时序关系或描述中间体形成与解离等动力学过程的要求。
静态单分子电学技术可以通过共价键构筑稳定连接的单个超分子组装体结构,具有较高的时间、电流分辨率和灵敏度,可以通过不同电流信号反映单个分子结动态结构变化,从而为纳米乃至亚纳米尺度下实时检测分子亚结构的动态变化提供了一个可靠的研究平台。通过采用上述技术构筑石墨烯-分子-石墨烯单分子结(GMG-SMJs),研究者已经实现了对单分子化学反应、立体电子效应、氢键组装体等的动力学研究。
一些光异构化分子,如偶氮衍生物、二苯基乙烯衍生物、螺旋藻衍生物等,光辐射可引起其构型的转变从而调节分子电导,是实现分子开关和逻辑门的有效方法。本工作通过上述GMG-SMJs器件,研究了1-[10-(4-苯基偶氮苯氧基)癸基]溴化吡啶(AzoC10)在紫外光-可见光下发生的异构化反应以及其光开关随机结合动力学。
如图1a所示,刚性共轭分子低聚苯乙炔(OPE3)通过酰胺共价键与石墨烯电极构筑成结,形成稳定的高导电通道,具有全甲基化侧基的α-环糊精(PM-α-CD)作为主体位于OPE3一侧。PM-α-CD与客体分子不同的结合状态会影响通道的电导。对α-环糊精(α-CD)进行全甲基化处理不仅能够防止脱水缩合过程中石墨烯电极边缘的羧基与α-CD的羟基发生反应,降低连接产率;而且能够破坏α-CD外表面的分子内氢键消除无关过程的干扰,提高信噪比。客体分子为两亲性AzoC10,由亲水性吡啶鎓单元、疏水性烷基链和偶氮苯三部分组成。如图1b所示,偶氮苯部分在紫外光照射下能够从反式转变为顺式,并在可见光照射从顺式转换回反式。由于CD空腔内表面与疏水客体之间的疏水和范德华相互作用,PM-α-CD可以很好地识别trans-AzoC10,而不能识别尺寸不匹配的cis-AzoC10。
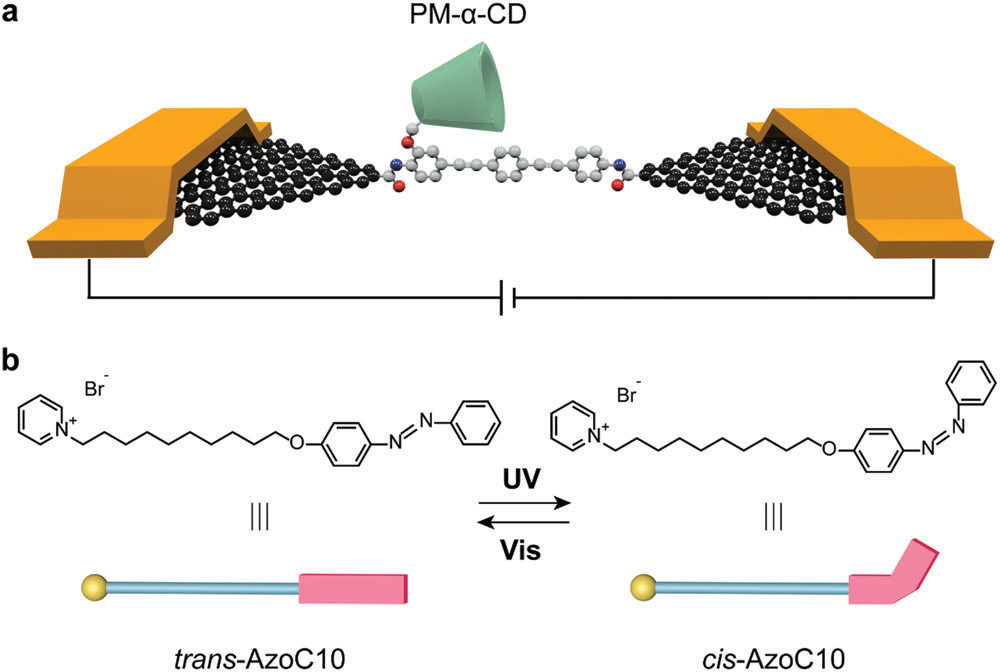
图1. 基于光切换的单个超分子组装体示意图。
(a)GMG-SMJs的器件结构。
(b)AzoC10的分子结构和示意图。黄色代表亲水吡啶鎓,蓝色代表疏水烷基链。红色代表偶氮苯部分,经紫外光和可见光的照射,偶氮苯部分在反式和顺式之间发生可逆光异构化反应。
测试前,在可见光(450 nm)或紫外光(365 nm)照射30分钟AzoC10水溶液,以确保分子完全处于反式或顺式。之后,在紫外光或可见光照射下进行实验,记录电流-时间(I-t)曲线,该I-t曲线直接反映了光辐射下超分子结随时间的结构变化。另外,测试过程中控制温度保持在303 K。
在可见光照射下,高斯拟合的电流直方图上显示出三个电流峰,表明trans-AzoC10在与环糊精空腔可逆结合和解离过程中存在三种不同的微观状态(图2a-b)。而当紫外光照射后,高电导状态消失,原始的三峰转变为双峰(图2c-d)。这可能由于偶氮苯由反式到顺式的转化。为了进一步验证光异构过程,在相同的器件上进行了连续可见光/紫外光照射的三循环电导测量实验。如图2e所示,在整个过程中,电流振荡信号I/I0(I-电流值,I0-每个周期的最低高斯拟合峰值)随切换光源在三种和两种信号状态之间循环变化,且每个状态下的I/I0值基本保持不变。上述变化的电导状态与AzoC10分子光异构化过程中与PM-α-CD空腔不同的结合构型对应。
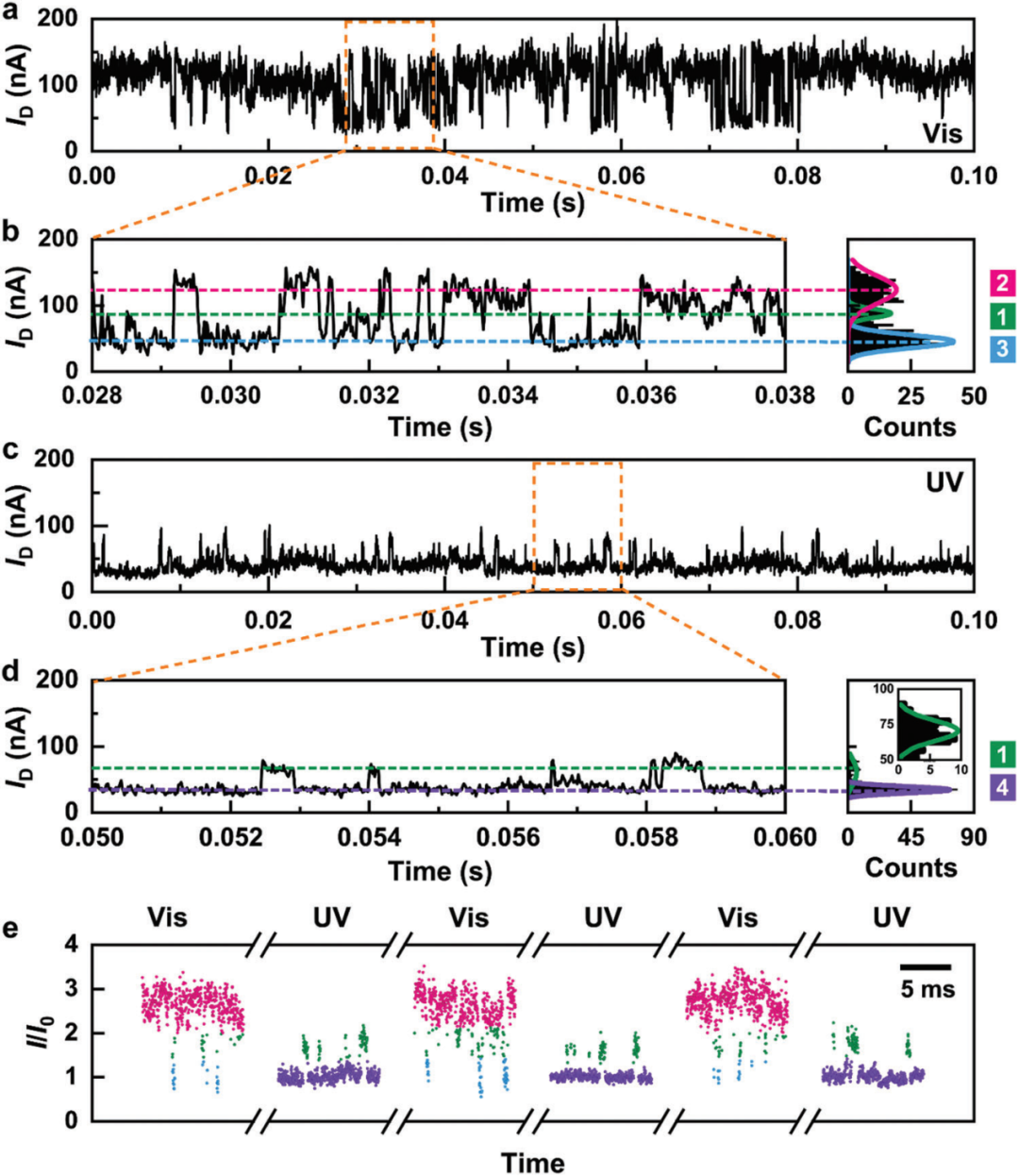
图2. 基于光切换的单个超分子组装体的原位实时电流-时间测试。
(a-d)303 K下1×10-8 M的AzoC10可逆结合过程0.1 s的I-t曲线及相应放大部分(a-b)可见光(450 nm)和(c-d)紫外线(365 nm)照射。
(b)和(d)中的右图是电流值的相应直方图,分别显示三峰和双峰电流分布。VD = 300 mV,采样率 = 57.6 kSa/s。
(e)连续的可见光和紫外光照射下三周期电流振荡信号(I/I0),颜色对应于(b)和(d)中的电流分布。
作者通过对分子电子结构和量子传输特性的理论计算,从而对Azoc10分子的各电导状态和对应的结合构型进行分析。如图3a所示,基于公式Ebinding = Gbinding-system – (Gmolecular + GPM-α-CD),作者对PM-α-CD与AzoC10分子的结合能进行了计算,结果表明反式偶氮苯和烷基链的结合能为负,而顺式偶氮苯和吡啶鎓单元的结合能为正。因此,前者可以被PM-α-CD很好地识别而后者不能。此外,反式偶氮苯的结合能低于烷基链,结合上述实验中状态2的分布在可见光下占据主导,而在紫外光下消失,作者认为状态2对应为反式偶氮苯与PM-α-CD的结合。而状态3的分布在可见光照下占中等比例,状态4的分布在紫外光下占据最高比例,分别对应于可见光和紫外光下烷基链与PM-α-CD的结合。状态1则对应PM-α-CD空腔。
此外,作者使用基于密度泛函理论的非平衡格林函数计算了不同分子构型的电荷传输特性(图3b-e),由于PM-α-CD两侧内径分别为0.45 nm和0.52 nm,AzoC10分子可以从两个方向与PM-α-CD结合,因此进行两种模式的计算,每种运动模式包含四种结合状态。两种模式下,p-HOMOs与电极费米能级的接近程度遵循相同的顺序,即反式偶氮苯、PM-α-CD空腔、顺式烷基链和反式烷基链,这也与实验测得各构型的电导顺序一致。
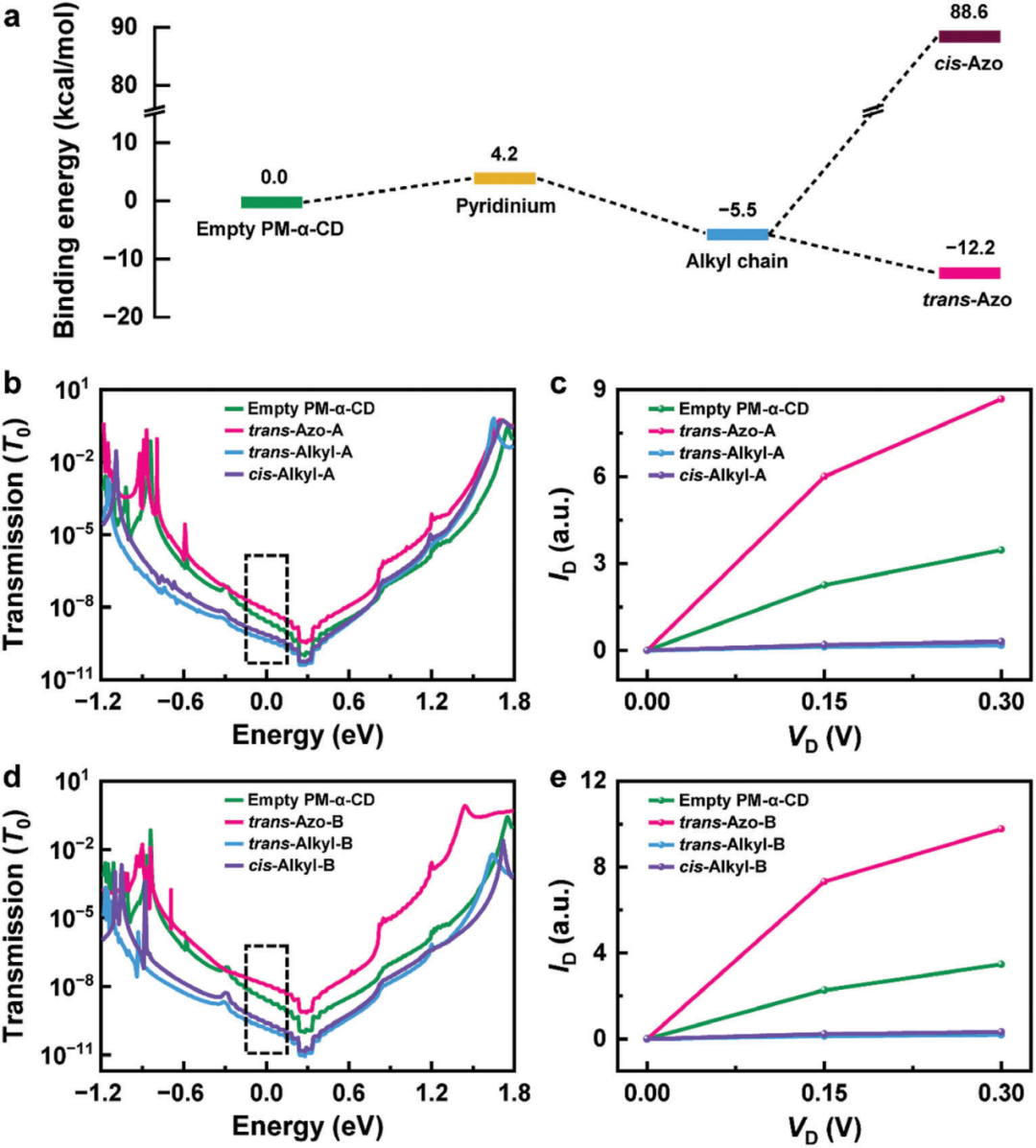
图3. 结合运动的理论分析。
(a)不同结合态的模拟能量分布。
在(b-c)构型A和(d-e)构型B中,GMG-SMJ在四种不同构型下的零偏置透射光谱和I-V曲线,(b)和(d)中的虚线框表示偏置窗口0.3 V。
根据电导状态的分配,作者对结合状态和异构体转换进行动力学分析。在可见光下,如图4a所示,反式构型呈现出五种典型的结合I-t曲线,分别对应三种往复运动过程和两种单向运动过程,其中前者出现的次数远大于后者。如图4b-g所示,基于马尔可夫模型对每个状态的停留时间进行单指数拟合,可以获得每个状态的平均寿命(τ)并计算出相应的速率常数(k = 1/τ)。得到的速率常数的大小关系k1→2 > k2→1, k1→3 > k3→1和k2→3 < k3→2 , 能够很好地对应反式偶氮苯、烷基链、PM-α-CD的热力学稳定性顺序。图4h表示的是整个结合过程的动力学模型。
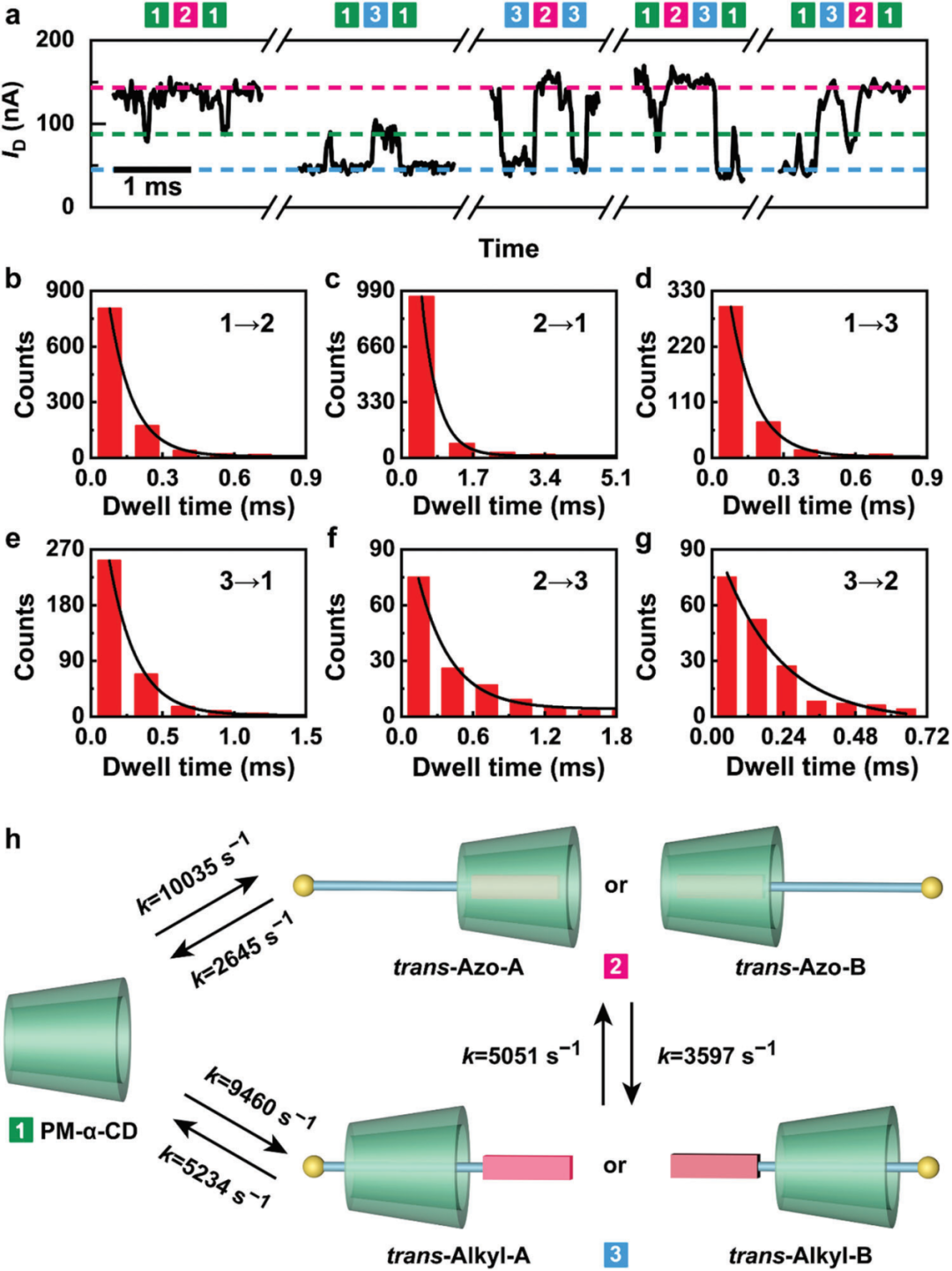
图4. 反式异构体的动力学分析。
(a)可见光照射下五种典型结合过程的I-t曲线。
(b)状态1→状态2,(c)状态2→状态1,(d)状态1→状态3,(e)状态3→状态1,(f)状态2→状态3和(g)状态3→状态2,停留时间统计。
(h)AB两种模式下结合过程的动力学模型。
伴随紫外光照射时间的增加,trans-AzoC10逐渐转化为cis-AzoC10。如图5所示,紫外光照5分钟后,作者观察到反式偶氮(状态2)的比例逐渐下降,反式烷基(状态3)和顺式烷基(状态4)共同存在较难区分。紫外光照30分钟后,反式偶氮(状态2)和反式烷基(状态3)完全消失,I-t曲线显示PM-α-CD空腔(状态1)和顺式烷基(状态4)的双峰分布。使用相同的方法对顺式构型进行动力学分析,如图5e-5g所示,得到的速率常数k同样与预期结果相符。
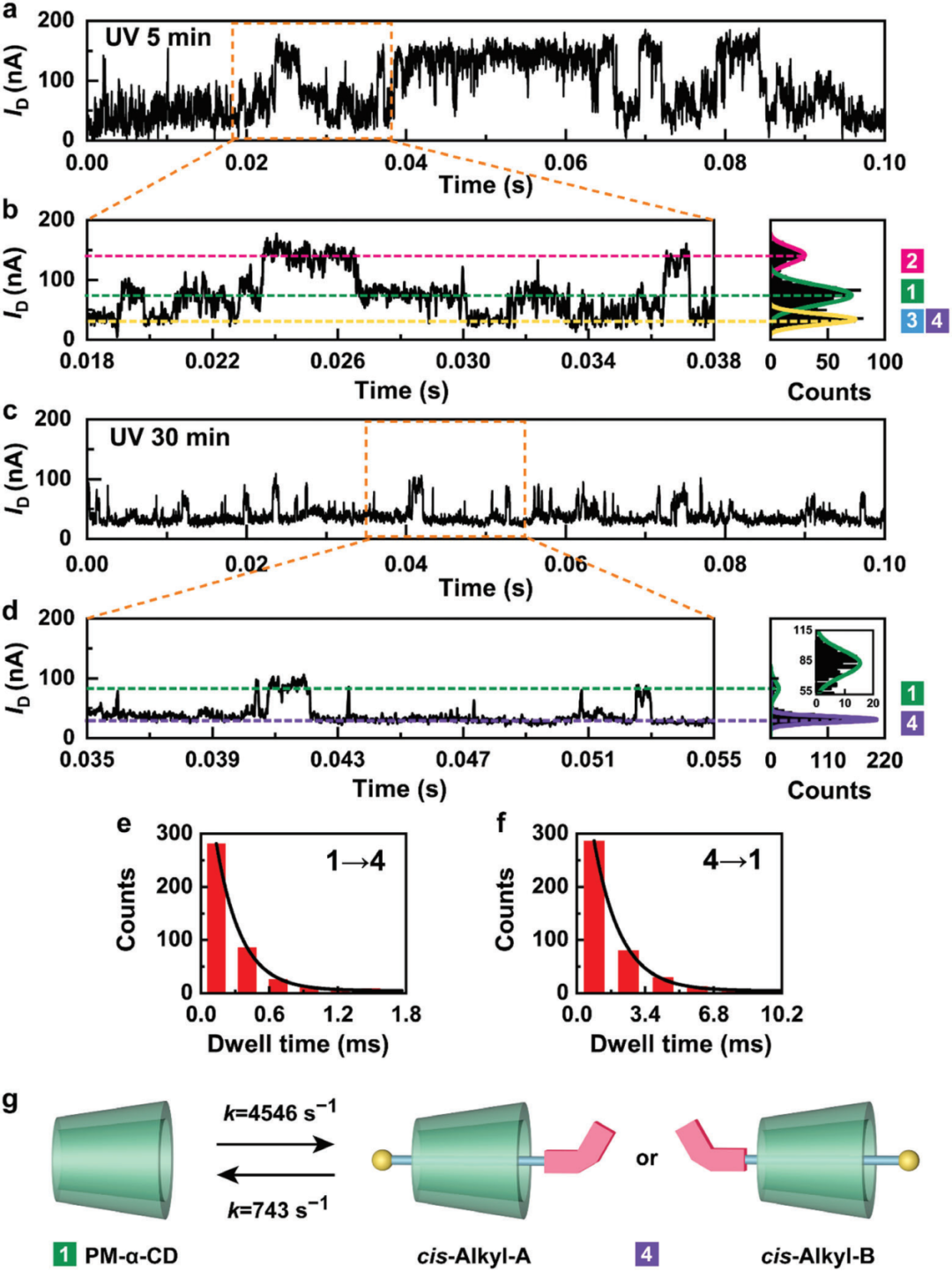
图5. 顺式异构体的动力学分析。
(a-b)5分钟和(c-d)30分钟紫外线照射下可逆结合过程的I-t曲线及相应的放大部分和电流分布的直方图。(e)状态1→状态4和(f)状态4→状态1,停留时间统计。
(g)结合过程的动力学模型。
本工作基于PM-α-CD和偶氮衍生物AzoC10之间的主客体相互作用,采用基于石墨烯固态电极的单分子电学方法实现了对单个超分子的随机运动的实时监测和动态表征。GMG-SMJs技术的高分辨率和灵敏度为从单分子水平直接表征AzoC10顺式构型和反式构型的不同微观运动模式及其在紫外光或可见光下的光异构化过程,并获得相应过程中的动力学参数提供了可能,对研究复杂系统的动态机制具有重要意义。此外,在客体分子上引入刺激响应功能单元的方法也为后续设计和制造新颖的单分子功能器件提供了新思路。
原文链接:https://onlinelibrary.wiley.com/doi/10.1002/advs.202200022
")
